The regulatory protein complex in Lethocerus and Drosophila flight muscle.
The
rapid oscillatory contraction of insect flight muscle is a consequence of
delayed activation of the muscle by stretch. A key property of the flight muscle is that, unlike other
muscles, it is not fully activated by calcium. Full activation only occurs when
the muscle is rapidly stretched. This year, we have made an important step
forward in understanding the mechanism of insect flight. It is now clear that
insect flight muscle contains two isoforms of troponin-C, which have different
regulatory functions. We have identified and sequenced the proteins in Lethocerus and have identified related TnC
sequences in the Drosophila and Anopheles genomes. TnC is the calcium regulated switch which activates the muscle during
contraction. We have measured the calcium binding of the insect TnCs by a
number of methods, including mass spectrometry (EMBL Proteomics Core Facility).
The
major isoform which makes up about 90% of TnC in the regulatory complex of the
flight muscle has only one calcium binding site and is not calcium sensitive.
The other, minor form, which comes
from a different gene, contains two calcium binding sites and regulates muscle
activity via calcium in the normal way
(fig 1). Stretch and
tension measurements on isolated
muscle fibres from Lethocerus from which the endogenous TnC was
removed and replaced by each of
the expressed isoforms in turn,
confirms that this is the case (work done in collaboration with W. Linke, Heidelberg).
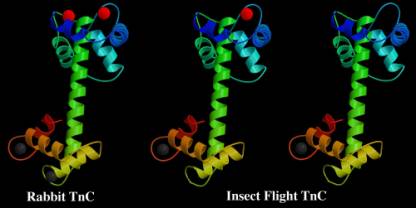
Fig.1 Vertebrate (rabbit) TnC on the left has four calcium ions bound per molecule. The grey calciums are strongly bound and do not exchange in vivo. The red calciums are less strongly bound and their presence or absence regulates the activity of the muscle. The insect flight muscle has two types of TnC. Both have only one strongly bound calcium (grey). The minor isoform has one exchangeable calcium (red) which can take part in normal activation. The major isoform has no exchangable calciums and would normally be inactive. This TnC is now thought to be the key to stretch activation.
High molecular weight
structural proteins and muscle elasticity
All striated muscles have large modular proteins (like
titin in vertebrates) which contribute to the elastic properties of the
muscle.The insect thorax contains muscles that vary widely in function,and the
ultra-structure of the sarcomere is correspondingly varied. The elastic
properties required of the different muscle fibres will determine which
titin-like proteins are present.The Drosophila D-titin
gene (annotated in Flybase as the SLS or ket gene)is predicted to code for a
protein similar to the N-terminal region of vertebrate titin.The maximum size
for a peptide from the Drosophila gene is
1.8 MDa,which is not large enough for the protein to extend from Z-disc to
M-line like vertebrate titin. Kettin, the shortest isoform (500 kDa), is
responsible for the unusually high stiffness of insect flight muscle. It is now
clear that related proteins exist in other invertebrates (fig 2).
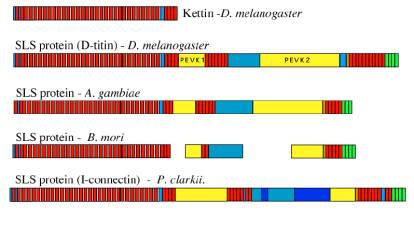
Fig 2.
Examples of
proteins of the SLS family. We have sequenced the most abundant isoform
in Drosophila flight muscle.which we have called kettin. Kettin is
made up of 35 Ig domains (red blocks) separated by 35 aa linkers and binds to
actin in the Z disk and I-band. Other, larger isoforms of the SLS proteins are
present in the extensible non-flight muscles of the insects but do not appear
from immunolabelling studies to extend further than from the Z-disk to the edge
of the A-band. Only partial sequences are available for the silk moth (B.
mori)
Key: red-Ig domains, green- Fn
domains, yellow-PEVK sequence, brown – SH3 domains, blue – undefined sequences
Amphipol stabilisation of Membrane Proteins
In collaboration with Jean-Luc Popot and Christophe Tribet (Paris) and Hanns Weiss, (Düsseldorf), we have been investigating the stability of the multi-subunit membrane protein complex NADH reductase (Complex I ) in the presence of amphipols with the goal of improving the structural information obtained by electron microscopy. The use of detergent for solubilising membrane proteins makes cryo-electron microscopy or lipid-monolayer crystallisation methods difficult or impossible owing to the excess of free detergent which lowers the surface tension. Amphipols are high molecular weight amphipholic polymers which bind tightly to the hydrophobic surfaces of the protein. Excess amphipol and detergent can then be removed resulting in solubilised protein without the problems caused by detergent.
W are using Complex I from Neurospora crassa mitochondria as a test object for this study. It is a very large integral membrane protein with a characteristic L-shape. One arm of the L is the hydrophobic membrane spanning domain which binds detergent and the other arm is the more hydrophilic cytoplasmic domain. The enzyme is first isolated from Neurospora crassa mitochondria and purified in either the detergent Triton X-100 or in dodecyl maltoside. The detergent is then exchanged with amphipol. We have concentrated our efforts on the effects of charged amphipol (A8-35) and neutral amphipol (A34-0) on the stability of Complex I. We have now made a preliminary cryo-EM 3-D reconstruction of CI with amphipol A8-35 which shows essentially the same structure as that previously obtained with negative stain. (Fig. 3)
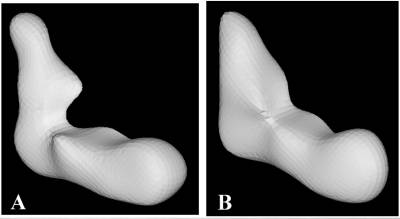
Fig 3.
(A) 3-D reconstruction of
Complex I made by the conical tilt method, form images of single particles
solubilised in detergent and stained negatively with uranyl acetate. The
horizontal arm is the membrane
domain and includes detergent. The vertical arm is the cytoplasmic
domain.
(B) 3-D reconstruction made by
the multi-reference alignment method for single particles stabilised (in the
absence of detergent) by amphipol A8-35. In this case, samples were unstained
and imaged frozen-hydrated. Both reconstructions were filtered to a cut-off of
3nm and at this resolution show comparable features.